XEPO BEN : LE CERVEAU A L'ESPACE A DEBATTRE 2 RUE VERNIER NICE
VERNISSAGE DE L'EXPOSITION
" PAS DE CERVEAU ...
... PAS DE CHOCOLAT ! "
AVEC BEN VAUTIER
à l'ESPACE à Débattre
2, Rue Vernier Nice
Samedi 13 Octobre 2012 à 18h30

Photo AGENCY PRESS INTERNATIONAL DIACONESCO.TV - C.COPYRIGHT - NICE 13.10.2012
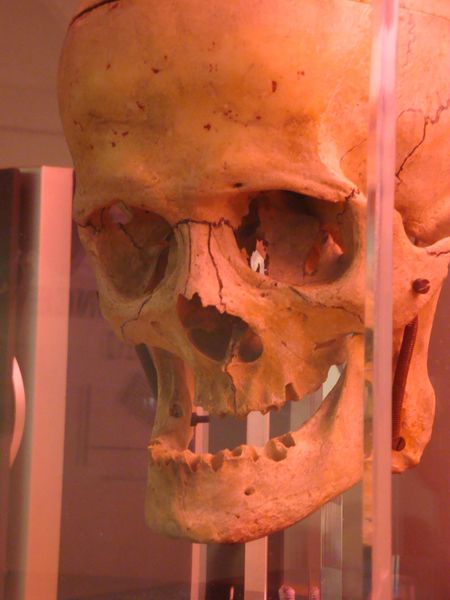
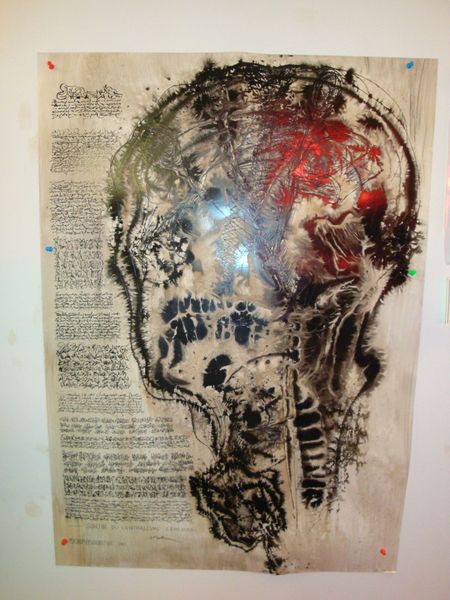
LE CERVEAU
avec BEN VAUTIER
2012-10-02
Bonjour,
Encore une newsletter de Ben avec 5 points à ne
pas oublier :
1ER POINT
Le repas du 3 octobre sur le ring :
à la Villa Arson
à partir de 18 h 33
ce sera « UNE AUBERGE ESPAGNOLE »
apportez votre pique nique.
fromages fruits etc.,
j’apporte à boire
et si vous y tenez
vous pouvez toujours réaliser
une performance elle sera filmée
2EME POINT
Important
J’arrête provisoirement les débats du vendredi
à 18h33 à l’Espace à débattre
La raison est simple : trop de monde mon cerveau
cale j’arrive pas à assumer j’aimerais peut-être un
jour remplacer les débats par des films-interviews
en solitaire des autopsies mentales.
3EME POINT
Important : l’expo sur LE CERVEAU
le 13 octobre
Pour l'expo sur le cerveau, j'attends vos
participations, c’est urgent, on accroche le
10 octobre, le vernissage c’est le 13 octobre.
J'ai déjà reçu la participation de Lizène. De ma
collection il y aura entre autres, Régent,
Robert Filliou, Combas, etc...
BIEN QUE LES MURS DE L’ESPACE A DEBATTRE
SOIENT PAS IMMENSES VOS IDEES, VOS DESSINS,
VOS TEXTES SERONT LES BIENVENUS AVANT LE 8
OCTOBRE SONT INVITES A PARTICIPER, S’ILS LE
VEULENT, A L’EXPO SUR LE « CERVEAU » :
ANNA BISKOV
ANTAKI
ARMLEDER JOHN
ARNAUD LABELLE-ROJOUX
ARNAUD MAGUET
BATAILLARD
BATLLE MICHEL
BEATRICE KUSSOL
BENICHOU
BOLTANSKI
BORIS CYRULNIK
BOUILLON
BRECHT GEORGE
BRUNO DUVAL
CATHERINE CATTANEO
CATHY GILBERT
CHARLES DREYFUS
COMBAS ROBERT
DALIGAND
DANIELE ROUX
DENIS CASTAGNOU
DUCKAERTS
EINSTEIN
FILLIOU
FRANÇOIS GUINOCHET
FRANÇOIS PARIS
FREDERIC ROUX
GERALD PANIGHI
GUICHOU
HELIOS AZOULAY
HENRY OLIVIER
JACQUES ET CATHERINE PINEAU
JACQUES LIZENE
JEAN BAPTISTE GANNE
JEAN DUPUY
JEAN JACQUES LEBEL
JEAN MAS
JEAN-CLAUDE GUILLAUMON
KARIM GELOUSSY
KOURO
KRIGLO
HUBAUT
ORLAN
LA GALLA
LA STATION
LEVY LEBLOND
LIONEL KOURAU
LIZENE JACQUES
LOUIS CANE
MAX HORDE
MAXIME MATRAY
MARCEL ALLOCO
MONIQUE THIBAUDIN
MOUTON JOSEPH
NICOLAS PRIVE
PHILIP CORNER
PHILIPPE LA HURE
PIERRE LE PILLOUËR
PIERRE TILMAN
PINAUD PASCAL
REGENT
DR ROBERT
RUY BLAS
SACHA SOSNO
SOPHIE CALLE
SOPHIE TAAN
SNYERS
WILLEM DE RIDDER
Etc...
LE TITRE DE L’EXPO
PAS DE CERVEAU
PAS DE CHOCOLAT !
POINT 4
AVIS IMPORTANT (LE FINISSAGE)
L’expo sur la performance de la villa Arson se termine avec un colloque important rempli de grosses têtes
(une performance en soi) les 26, 27, 28 octobre si je ne suis pas malade (du cerveau) et que ma mémoire ne flanche pas j’aimerai ouvrir le jeudi 26 octobre le ring avec une soirée de nouvelles performances de 3 minutes 33 un genre de
FINISSAGE QUE CEUX QUI VEULENT PARTICIPER
le jeudi 26 à partir de 18h33 me le communiquent par contre avis important il n’y a plus d’argent disponible sur les 5.000 Euros de budget j’ai dépassé les 9.000 Euros !
POINT 5
Je vais demander à Benoit de me faire une vidéo réduisant les
3 minutes 33 en 30 secondes 33 pour chaque performance
vu que c’est impossible de voir le tout plus de 16 heures
COMPTE RENDU DE CE QUI S’EST PASSÉ
MERCREDI 26 SEPTEMBRE 2012 SUR LE RING
Je m’attendais comme à la bourse à une séance morose la surprise vint de l’énergie des participants et l’ambiance d’un feeling de liberté de création
Un très bon Erebo qui filme les pieds et le plafond
J'ai bien aimé la pièce de la farine
J'ai bien aimé la pièce de Cataneo
J’ai bien aimé Sophie Taan et sa pièce sur Darwin
J’ai bien aimé le chant de Setsuko,
japonaise venant d’Italie, très émouvant
et j’ai même pu supporter Lita Chow
son ego sous son papier aluminium…
etc...
VENDREDI DERNIER AU DEBAT DE
18H33 A L’ESPACE A DEBATTRE
Débat sur le cerveau
Ça partait dans tous les sens, et pourtant, soudain je me suis rendu compte, que la terre est composée de 7 milliards de cerveaux tous à peu de chose près identiques,
En poids et en taille avec tous le même moteur, un ego qui cherche à survivre.
En dehors de ces 7 milliards de cerveaux,
Sur le plan explication de l’univers il n'y a presque rien qui existe.
Donc, on peut imaginer le monde, mu par un engrenage de deux milliards de cerveaux qui s'intercalent, chaque cerveau contenant des milliards de neurones.
THEORIE
La prochaine révolution, qui pourrait changer, les rapports
entres les groupes ethniques entre femmes et hommes, serait La télépathie : savoir ce que les autres pensent.
IDEE FILM
Une série de plans fixes,
x tient à la main un petit appareil
une machine à lire les pensées.
On filme les pensées d’une belle femme
On entend deux pensées,
la pensée de ce que nous croyons
que l’actrice pense
et la pensée de celui qui filme.
BEN CINEASTE
Le film 48h est en marche,
J’ai appris à faire les textes
et à intercaler les images.
BEN THEORICIEN
J’ignore si la diversité
(la diversification)
gagne ou perd du terrain
sur l'extinction (de la vie)
Y a t'il plus d'espèces ou moins d'espèces ?
Deuxième question
la diversité est-elle un plus pour l'espèce humaine?
Autre question
Faut il être fataliste et se réjouir
Quand il y a extinction ?
C'est la loi du plus fort....donc
c'est normal qu'il y ait disparition.
Un peu confus tout cela
BEN THEORICIEN
Bonjour
important
je me regarde dans le miroir
qui au juste es-tu ?
difficile à cerner.
NICE CULTURE
Yves Hayat chez Robert Roux
Le 18 octobre
116 bld de la Madeleine
titre de l’expo :
« The business must go on »
Des Photos de villes en ruines
Où seuls survivent dans les ruines
Coca cola
Mac Donald etc
Yves est notre grand peintre
attaché au surréalisme et au beau.
NICE CULTURE
Catherine Issert est partie pour Paris
pour sélectionner les galeries de ART/PARIS.
Je lui demande s’il y aura des galeries Niçoises,
elle a dit : non je ne le crois pas à part moi.
BEN THEORICIEN
Mon livre ''tout est ego'' est en marche,
mais je ne l'aime pas,
je suis ridicule
je pontifie inutilement
grosse déception
je suis lucide
ce n’est pas de la théorie
ce n’est que de la prétention
je théorise tout.
Il y a eu une petite bifurcation
et l’espèce humaine est devenue prédatrice
si j’efface l’ego écrit sur le tableau noir
l’ego disparaît-il aussi ?
Est-ce parce qu’il n’y a plus d’ego pour voir le tableau
ou est-ce parce que le tableau n’a jamais existé ?
BEN THEORICIEN
Je ne relis pas mes anciens textes
et quand je les relis
je ne sais pas quoi en penser
Je ne sais pas si
c’est une bonne ou une mauvaise chose
alors je garde tout sur mon ordinateur
tant pis pour moi si je suis ridicule
je préfère être ridicule qu’absent.
BEN THEORICIEN
Je crois sincèrement que si je pouvais
faire disparaître l’espèce humaine
avec une baguette magique
je le ferais.
BEN SUR BEN
Important pour moi
De dire le contraire
De me lâcher
De déplaire
De me taire
De me montrer nu
De me cacher
De mentir
De ne pas croire en l’art
De m’habiller à l’abbé Pierre
De raconter des bêtises
De me sentir heureux
De cracher dans la soupe
BEN » TOUT EST EGO «
Il est temps de désamorcer ce livre
Que des conneries
Aucune épine dorsale
Aucune théorie digne de ce nom
La plus simple des revues de sciences
contient dix fois plus
Et pourtant je persiste
Pour théoriser il suffit de supposer
supposons que rien n’existe
Supposons que tout est différent
Supposons le contraire de tout
Fin de ma théorie des supposons
je gaspille trop de papier
la nature se vengera un jour sur moi
BEN ET CAGE
Eric Mangion me demande l’autorisation
de distribuer à la Sorbonne mon tract contre Cage
je suis ni pour ni contre.
LA CRISE ET L'ART
Un critique d’art m’a dit :
Dévaluation ou pas
Chômage ou pas de chômage,
Une oeuvre restera dans le temps
toujours unique
donc quand on tient à la main,
une pièce unique
on peut se dire ''crise ou pas crise''
si cette pièce apporte du nouveau
je ferais mieux de la garder ou
de la vendre.
IL FAUT QUE JE VOUS DISE QUE
pour Blois
ça avance
la cour est presque finie
il va y avoir des questions partout dans la ville
il va y avoir la collection de Gino
ma collection à moi aussi
l’important est que le bar marche
avec de belles serveuses
et qu’il y ait une bonne bibliothèque
avec tous les livres sur Fluxus et la
performance.
DANS LE MONDE
L’intolérance double
l’hypocrisie règne
Aujourd’hui
Le cinéma autorise de
se mettre à la place d’un extra terrestre
qui arrive et dit :
L’espèce humaine est fantastique
mais elle doit disparaître.
NICE CULTURE
Le Nissart occitan
Gagne du terrain
De plus en plus de chanteurs de groupes
La place Garibaldi ;
une grande réussite
positif entre autre parce que
si la crise s’aggrave
La solution a la crise économique
Sera de compter sur le dynamisme local
Donc
Initiative Innovation création
NICE CULTURE
Cataneo veut réaliser un film
sur la vie d’Olga Adorno
née en 1937
Performeuse dans le pop art et
comme Ultra Violet
une grande muse.
BEN SUR BEN
C’est compliqué je navigue sans boussole
perdu dans l’océan
Le portail ne s’ouvrait pas il s’ouvre maintenant
Il y a de la poussière sur tous les tableaux
L’expo Catherine Issert ne devrait pas
me prendre la tête et pourtant j’ai eu de l’angoisse
j’en ai marre de mon égo
A Wiesbaden ils m’attendaient
Mais grève des avions Lufthansa.
POLITIQUE EXTERIEURE FRANCAISE
ERREUR EN LYBIE
(impérialisme inutile)
la preuve : on est en train d’en payer les conséquences
la situation intérieure est peut-être
pire que du temps de Kadhafi
ERREUR AU MALI
Attention au bourbier
les peuples existent
au Mali où la résistance des Berbères Touareg
veut se libérer du pouvoir central de Bamako
la France veut garder l’uranium du Niger d’Areva.
Il ne faut pas oublier que le sud est Dogon et Bambara
le nord du Mali est Touareg
il faut arrêter cette comédie « d’intégrité du territoire »
ERREUR EN SYRIE
Trop de confusion géo-stratégique
trop grand risque d’engrenage
Le rapprochement des Kurdes de Syrie
avec le PKK,
est d’après moi positif pour les Kurdes
mais négatif pour les Turcs
(impérialistes envers les Kurdes )
Conséquence de cette guerre
Difficile de faire croire que
Que les 80 millions d’Egyptiens
Vont rester là les bras croisés à laisser
le Qattar décider de tout
LE MONDE
La Belgique
se divise calmement,
sans trop de morts.
Si seulement, cela pouvait être
la même chose pour le Soudan,
le Tibet, la Côte d’Ivoire
le Kurdistan ; etc.
Important
pour mes nouveaux abonnés sachez
que vous pouvez lire mes anciennes newsletter sur mon site
http://www.ben-vautier.com/
à la rubrique « mes newsletter »
Ben
LES PHOTOS DE LA SOIREE AVEC BEN
C.COPYRIGHTS ALL RIGHTS AGENCY PRESS INTERNATIONAL
DISCONESCO.TV - NICE 13.10.2012






























































PHOTOS C. COPYRIGHTS AGENCY PRESS INTERNATIONAL DIACONESCO.TV - NICE 13.10.2012
SUR WIKIPEDIA DICTIONNAIRE
Le Cerveau
 Pour l’article homophone, voir Servo.
Pour l’article homophone, voir Servo.Le cerveau est le principal organe du système nerveux des animaux. Au sens strict, le cerveau est l'ensemble des structures nerveuses dérivant du prosencéphale (diencéphale et télencéphale). Dans le langage courant, ce terme peut désigner soit l'encéphale dans son ensemble, soit le seul télencéphale ou même le seul cortex cérébral.
Chez les vertébrés, le cerveau est situé dans la tête, protégé par le crâne, et son volume varie grandement d'une espèce à l'autre. Par analogie, chez les invertébrés, le cerveau désigne certains centres nerveux.
Le cerveau régule les autres systèmes d'organes du corps, en agissant sur les muscles ou les glandes, et constitue le siège des fonctions cognitives. Ce contrôle centralisé de l'organisme permet des réponses rapides et coordonnées aux variations environnementales. Les réflexes, schémas de réponses simples, ne nécessitent pas l'intervention du cerveau. Toutefois, les comportements plus sophistiqués nécessitent que le cerveau intègre les informations transmises par les systèmes sensoriels et fournissent une réponse adaptée.
Le cerveau est une structure extrêmement complexe qui peut renfermer jusqu'à plusieurs milliards de neurones connectés les uns aux autres. Les neurones sont les cellules cérébrales qui communiquent entre elles par le biais de longues fibres protoplasmiques appelées axones. L'axone d'un neurone transmet des influx nerveux, les potentiels d'action, à des cellules cibles spécifiques situées dans des régions plus ou moins distantes du cerveau ou de l'organisme. Les cellules gliales sont le deuxième type cellulaire du cerveau et assurent des fonctions très diversifiées, centrées autour du support des neurones et de leurs fonctions.
Malgré de grandes avancées en neurosciences, le fonctionnement du cerveau est encore mal connu. Les relations qu'il entretient avec l'esprit sont le sujet de nombreuses discussions, aussi bien philosophiques que scientifiques.
Anatomie

Schéma d'organisation fondamental d'un bilatérien.
Le cerveau est la structure biologique la plus complexe connue1 ce qui rend souvent délicate la comparaison de cerveaux de différentes espèces à partir de leur apparence. Néanmoins, l'architecture du cerveau présente plusieurs caractéristiques communes à un grand nombre d'espèces. Trois approches complémentaires permettent de les mettre en évidence. L'approche évolutionniste compare l'anatomie du cerveau entre différentes espèces et repose sur le principe que les caractères retrouvés sur toutes les branches descendantes d'un ancêtre donné étaient aussi présentes chez leur ancêtre commun. L'approche développementale étudie le processus de formation du cerveau du stade embryonnaire au stade adulte. Enfin, l'approche génétique analyse l'expression des gènes dans les différentes zones du cerveau.
L'origine du cerveau remonte à l'apparition des bilatériens, une des principales subdivisions du règne animal notamment caractérisée par une symétrie bilatérale des organismes, il y a environ 550-560 millions d'années2. L'ancêtre commun de ce taxon suivait un plan d'organisation de type tubulaire, vermiforme et métamérisé ; un schéma qui continue de se retrouver dans le corps de tous les bilatériens actuels, dont l'Homme3. Ce plan d'organisation fondamental du corps est un tube renfermant un tube digestif, reliant la bouche et l'anus, et un cordon nerveux qui porte un ganglion au niveau de chaque métamère du corps et notamment un ganglion plus important au niveau du front appelé « cerveau ».
Invertébrés
La composition du cerveau des invertébrés est très différente de celle des vertébrés, à tel point qu'il est difficile de comparer les deux structures à moins de se baser sur la génétique. Deux groupes d'invertébrés se démarquent par un cerveau relativement complexe : les arthropodes et les céphalopodes4. Le cerveau de ces deux groupes provient de deux cordons nerveux parallèles qui s'étendent à travers tout le corps de l'animal. Les arthropodes ont un cerveau central avec trois divisions et de larges lobes optiques derrière chaque œil pour le traitement visuel4. Les céphalopodes possèdent le plus gros cerveau de tous les invertébrés. Le cerveau des pieuvres est très développé, avec une complexité similaire à celle rencontrée chez les vertébrés.
Le cerveau de quelques invertébrés a été particulièrement étudié. Par la simplicité et l'accessibilité de son système nerveux, l'aplysie a été choisie comme modèle par le neurophysiologiste Eric Kandel pour l'étude des bases moléculaires de la mémoire qui lui valut un Prix Nobel en 20005. Cependant, les cerveaux d'invertébrés les plus étudiés demeurent ceux de la drosophile et du ver nématode Caenorhabditis elegans. Du fait de l'important panel de techniques à disposition pour étudier leur matériel génétique, les drosophiles sont tout naturellement devenues un sujet d'étude sur le rôle des gènes dans le développement du cerveau6. De nombreux aspects de la neurogénétique des drosophiles se sont avéré être également valable chez l'Homme. Par exemple, les premiers gènes impliqués dans l'horloge biologique furent identifiés dans les années 1970 en étudiant des drosophiles mutantes montrant des perturbations dans leur cycles journaliers d'activité7. Une recherche sur le génome des vertébrés a montré un ensemble de gènes analogues à ceux de la drosophile jouant un rôle similaire dans l'horloge biologique de la souris et probablement également dans celle de l'Homme8. Comme la drosophile, le nématode C. elegans a été très étudié en génétique9 car son plan d'organisation est très stéréotypé : le système nerveux du morphe hermaphrodite possède exactement 302 neurones, toujours à la même place, établissant les mêmes liaisons synaptiques pour chaque ver10. Au début des années 1970, du fait de sa simplicité et de sa facilité d’élevage, Sydney Brenner le choisit comme organisme modèle pour ses travaux sur le processus de régulation génétique du développement qui lui valurent un Prix Nobel en 200211. Pour ses travaux, Brenner et son équipe ont découpé les vers en milliers de sections ultra fines et photographié chacune d'entre elles au microscope électronique afin de visualiser les fibres assorties à chaque section et ainsi planifier chaque neurone et chaque synapse dans le corps du ver12. Actuellement, un tel niveau de détail n'est disponible pour aucun autre organisme, et les informations récoltées ont rendu possibles de nombreuses études.
Vertébrés

Comparaison des cerveaux de différentes espèces
Apparus il y a 500 millions d'années, les vertébrés ont dérivé d'une forme proche de la myxine actuelle13. Le cerveau de tous les vertébrés présente fondamentalement la même structure14. Au cours de l'évolution de cet embranchement, la tendance évolutive du cerveau a été de suivre un gradient de taille et de complexité croissante, en particulier chez les mammifères15.
Le cerveau des vertébrés est d'un tissu mou et d'une texture gélatineuse16. De manière générale, le tissu cérébral vivant est rosâtre à l'extérieur et blanchâtre à l'intérieur. Le cerveau des vertébrés est enveloppé d'un système membranaire de tissu conjonctif, les méninges, qui sépare le crâne du cerveau17. De l'extérieur vers l'intérieur, les méninges sont composées de trois membranes : la dure-mère, l'arachnoïde et la pie-mère. L'arachnoïde et la pie-mère sont étroitement connectées entre elles et peuvent ainsi être considérées comme une seule et même couche, la pie-arachnoïde. Compris entre l'arachnoïde et la pie mère, l'espace sous-arachnoïdien contient le liquide cérébro-spinal qui circule dans l'étroit espace entre les cellules et à travers les cavités appelées système ventriculaire. Ce liquide sert notamment de protection mécanique au cerveau en absorbant et amortissant les chocs et à transporter hormones et nutriments vers le tissu cérébral. Les vaisseaux sanguins viennent irriguer le système nerveux central à travers l'espace périvasculaire au-dessus de la pie-mère. Au niveau des vaisseaux sanguins, les cellules sont étroitement jointes, formant la barrière hémato-encéphalique qui protège le cerveau en agissant comme un filtre vis-à-vis des toxines susceptibles d'être contenues dans le sang.
Tous les cerveaux des vertébrés possèdent la même forme sous-jacente observable par la manière dont le cerveau se développe18. Pendant le neurodéveloppement, le système nerveux commence à se mettre en place par l'apparition d'une fine bande de tissu neural parcourant tout le dos de l'embryon. La bande s'épaissit ensuite et se plisse pour former le tube neural. C'est à l'extrémité crânienne du tube que se développe le cerveau, l'émergence de ce crâne chez les premiers vertébrés aquatiques étant en relation avec le développement de leur sens de l'olfaction lié à leur capacités exploratrices à la recherche de proies. Au départ, le cerveau se manifeste comme trois gonflements qui représentent en fait le prosencéphale, le mésencéphale et le rhombencéphale. Chez de nombreux groupes de vertébrés, ces trois régions gardent la même taille chez l'adulte, mais le prosencéphale des mammifères devient plus important que les autres régions et le mésencéphale plus petit19.
La relation entre la taille du cerveau, la taille de l'organisme et d'autres facteurs a été étudiée à travers un grand nombre d'espèces de vertébrés. La taille du cerveau augmente avec la taille de l'organisme, mais pas de manière proportionnelle. Chez les mammifères, la relation suit une loi de puissance, avec un exposant d'environ 0,7520. Cette formule s'applique pour le cerveau moyen des mammifères mais chaque famille s'en démarque plus ou moins, reflétant la complexité de leur comportement. Ainsi, les primates ont un cerveau cinq à dix fois plus gros que ce qu’indique la formule. De manière générale, les prédateurs tendent à avoir des cerveaux plus gros. Quand le cerveau des mammifères augmente en taille, toutes les parties n'augmentent pas dans la même proportion. Plus le cerveau d'une espèce est gros, plus la fraction occupée par le cortex est importante21, 80% de l'activité cérébrale dépendant des signaux visuels chez les primates19.
Régions du cerveau
En neuroanatomie des vertébrés, le cerveau est généralement considéré comme constitué de six régions principales définies sur la base du développement du système nerveux à partir du tube neural : le télencéphale, le diencéphale, le mésencéphale, le cervelet, le pont, et le bulbe rachidien22. Chacune de ces régions possède une structure interne complexe. Certaines régions du cerveau, comme le cortex cérébral ou le cervelet, sont formés de couches formant des replis sinueux, les circonvolutions cérébrales, qui permettent d'augmenter la surface corticale tout en logeant dans la boîte crânienne. Les autres régions du cerveau représentent des groupes de nombreux noyaux. Si des distinctions claires peuvent être établies à partir de la structure neurale, la chimie et la connectivité, des milliers de régions distinctes peuvent être identifiées dans le cerveau des vertébrés.
Chez plusieurs branches des vertébrés, l'évolution a amené des changements importants sur l'architecture du cerveau. Les composants du cerveau des requins sont assemblés de façon simple et directe, mais chez les poissons téléostéens, groupe majoritaire des poissons modernes, le prosencéphale est devenu éverté. Le cerveau des oiseaux présente également d'importants changements23. Un des principaux composants du prosencéphale des oiseaux, la crête ventriculaire dorsale, a longtemps été considéré comme l'équivalent du ganglion basal des mammifères, mais est maintenant considéré comme étroitement apparenté au néocortex24.
De nombreuses régions du cerveau ont gardé les mêmes propriétés chez tous les vertébrés1. Le rôle de la plupart de ces régions est encore soumis à la discussion mais il est malgré tout possible de dresser une liste des régions principales du cerveau et le rôle qu'on leur attribue selon les connaissances actuelles :

Les principales divisions de l'encéphale représentées sur un cerveau de requin et un cerveau humain
- Le bulbe rachidien (ou medulla oblongata) prolonge la moelle épinière. Elle contient de nombreux petits noyaux impliqués dans un grand nombre de fonctions motrices et sensitives25.
- L'hypothalamus est un petit organe situé sous le prosencéphale. Il est composé de nombreux petits noyaux possédant chacun ses propres connexions et une neurochimie particulière. L'hypothalamus régule et contrôle de nombreuses fonctions biologiques essentielles telles que l'éveil et le sommeil, la faim et la soif, ou la libération d'hormones26.
- Le thalamus est également composé de noyaux aux fonctions diverses. Une partie d'entre eux servent à relayer l'information entre les hémisphères cérébraux et le tronc cérébral. D'autres sont impliqués dans la motivation. La zona incerta, ou région sous-thalamique, semble jouer un rôle dans plusieurs comportements élémentaires comme la faim, la soif, la défécation et la copulation27.
- Le cervelet joue un rôle majeur dans la coordination des mouvements en modulant et optimisant les informations provenant d'autres régions cérébrales afin de les rendre plus précises. Cette précision n'est pas acquise à la naissance et s'apprend avec l'expérience28.
- Le tectum, partie supérieure du mésencéphale, permet de diriger les actions dans l'espace et de conduire le mouvement. Chez les mammifères, l'aire du tectum la plus étudiée est le colliculus supérieur qui s'occupe de diriger le mouvement des yeux. Le tectum reçoit de nombreuses informations visuelles, mais aussi les informations d'autres sens qui peuvent être utiles pour diriger les actions comme l'ouïe. Chez certains poissons, comme la lamproie, le tectum occupe la plus large partie du cerveau29.
- Le pallium est une couche de matière grise qui s'étale sur la surface du prosencéphale. Chez les mammifères et les reptiles, il est appelé cortex cérébral. Le pallium est impliqué dans de nombreuses fonctions telles que l'olfaction et la mémoire spatiale. Chez les mammifères, il s'agit de la région dominante du cerveau et elle subsume les fonctions de nombreuses régions sous-corticales30.
- L'hippocampe, au sens strict, n'est présent que chez les mammifères. Néanmoins, cette région dérive du pallium médial communs à tous les vertébrés. Sa fonction est encore mal connue mais cette partie du cerveau intervient dans la mémoire spatiale et la navigation31.
- Les ganglions de la base sont un groupe de structures interconnectées situées dans le prosencéphale. La fonction principale de ces ganglions semble être la sélection de l'action. Ils envoient des signaux inhibiteurs à toutes les parties du cerveau qui peuvent générer des actions et, dans les bonnes circonstances, peuvent lever l'inhibition afin de débloquer le processus et permettre l'exécution de l'action. Les récompenses et les punitions exercent, au niveau des ganglions de la base, des effets neurologiques conséquents sur ce dernier32.
- Le bulbe olfactif est une structure particulière qui traite les signaux olfactifs et envoie l'information vers la zone olfactive du pallium. Chez beaucoup de vertébrés, le bulbe olfactif est très développé mais il est plutôt réduit chez les Primates33.
Mammifères
Le cortex cérébral est la région du cerveau qui distingue au mieux le cerveau des Mammifères de celui des autres Vertébrés, celui des Primates de celui des autres Mammifères, et celui des Hommes de celui des autres Primates. Le rhombencéphale et le mésencéphale des Mammifères est généralement similaire à celui des autres vertébrés, mais des différences très importantes se manifestent au niveau du prosencéphale qui n'est pas seulement beaucoup plus gros mais présente également des modifications dans sa structure34. Chez les autres vertébrés, la surface du télencéphale est recouverte d'une simple couche, le pallium35. Chez les Mammifères, le pallium a évolué en une couche à six feuillets appelée néocortex. Chez les Primates, le néocortex s'est grandement élargi, notamment au niveau de la région des lobes frontaux. L'hippocampe des mammifères a également une structure bien particulière.
L'histoire évolutive de ces particularités mammaliennes, notamment le néocortex, est difficile à retracer35. Les synapsides, ancêtres des Mammifères, se sont séparés des sauropsides, ancêtres des reptiles actuels et des oiseaux, il y a environ 350 millions d'années. Ensuite, il y a 120 millions d'années, les mammifères se sont ramifiés en monotrèmes, marsupiaux et placentaires, division qui a abouti aux représentants actuels. Le cerveau des monotrèmes et des marsupiaux se distingue de celui des placentaires (groupe majoritaire des Mammifères actuels) à différents niveaux, mais la structure de leur cortex cérébral et de leur hippocampe est la même. Ces structures ont donc probablement évolué entre -350 et -120 millions d'années, une période qui ne peut être étudiée qu'à travers les fossiles mais ceux-ci ne préservent pas les tissus mous comme le cerveau.
Primates
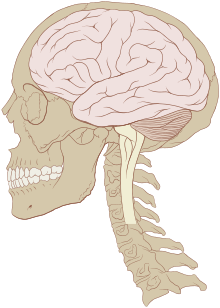
Schéma d'un cerveau humain dans sa boite crânienne.
Le cerveau des primates possède la même structure que celui des autres mammifères, mais il est considérablement plus large proportionnellement à la taille de l'organisme21. Cet élargissement provient essentiellement de l'expansion massive du cortex, notamment au niveau des régions servant à la vision et à la prévoyance36. Le processus de perception visuelle chez les Primates est très complexe, faisant intervenir au moins trente zones distinctes et un important réseau d'interconnexions, et occupe plus de la moitié du néorcortex37. L'élargissement du cerveau provient également de l'élargissement du cortex préfrontal dont les fonctions sont difficilement résumables mais portent sur la planification, la mémoire de travail, la motivation, l'attention, et les fonctions exécutives.
Chez les humains, l'élargissement des lobes frontaux est encore plus extrême, et d'autres parties du cortex sont également devenues plus larges et complexes.
Histologie
Le tissu cérébral est composé de deux types de cellules, les neurones et les cellules gliales38. Les neurones jouent un rôle prépondérant dans le traitement de l'information nerveuse tandis que les cellules gliales, ou cellules de soutien, assurent diverses fonctions annexes dont le métabolisme cérébral. Bien que ces deux types de cellules soient en même quantité dans le cerveau, les cellules gliales sont quatre fois plus nombreuses que les neurones dans le cortex cérébral39.
Contrairement aux cellules gliales, les neurones sont capables de communiquer entre eux à travers de longues distances40. Cette communication se fait par des signaux envoyés par le biais de l'axone, prolongement protoplasmique du neurone qui s'étend depuis le corps cellulaire, se ramifie et se projette, parfois vers des zones proches, parfois vers des régions plus éloignées du cerveau ou du corps. Le prolongement de l'axone peut être considérable chez certains neurones. Les signaux transmis par l'axone se font sous forme d'influx électrochimiques, appelés potentiels d'action, qui durent moins d'un millième de seconde et traversent l'axone à une vitesse de 1 à 100 mètres par seconde. Certains neurones émettent en permanence des potentiels d'action, de 10 à 100 par seconde, d'autres n'émettent des potentiels d'action qu'occasionnellement.
Le point de jonction entre l'axone d'un neurone et un autre neurone, ou une cellule non-neuronale, est la synapse où le signal est transmis41. Un axone peut avoir jusqu'à plusieurs milliers de terminaisons synaptiques. Lorsque le potentiel d'action, après avoir parcouru l'axone, parvient à la synapse, cela provoque la libération d'un agent chimique appelé neurotransmetteur. Une fois libéré, le neurotransmetteur se lie aux récepteurs membranaires de la cellule cible. Certains récepteurs neuronaux sont excitateurs, c'est-à-dire qu'ils augmentent la fréquence de potentiel d'action au sein de la cellule cible ; d'autres récepteurs sont inhibiteurs et diminuent la fréquence de potentiel d'action ; d'autres ont des effets modulatoires complexes.
Les axones occupent la majeure partie de l'espace cérébral42. Les axones sont souvent regroupés en larges groupes pour former des faisceaux de fibres nerveuses. De nombreux axones sont enveloppés d'une gaine de myéline, une substance qui permet d'augmenter fortement la vitesse de propagation du potentiel d'action. La myéline est de couleur blanche, de telle sorte que les régions du cerveau essentiellement occupées par ces fibres nerveuses apparaissent comme de la substance blanche tandis que les zones densément peuplées par les corps cellulaires des neurones apparaissent comme de la substance grise. La longueur totale des axones myélinisés dans le cerveau adulte d'un Humain dépasse en moyenne les 100 000 kilomètres43.
Développement

Principales subdivisions du cerveau embryonnaire des Vertébrés.
Le développement du cerveau suit une succession d'étapes44. Beaucoup de neurones naissent dans des zones spécifiques contenant des cellules souches et migrent ensuite à travers le tissu pour atteindre leur destination ultime45. Ainsi, dans le cortex, la première étape du développement est la mise en place d'une armature par un type de cellules gliales, les cellules radiales, qui établissent des fibres verticales à travers le cortex. Les nouveaux neurones corticaux sont créés à la base du cortex et « grimpent » ensuite le long des fibres radiales jusqu'à atteindre les couches qu'ils sont destinés à occuper.
Chez les vertébrés, les premières étapes du développement sont communes à toutes les espèces44. Tandis que l'embryon passe d'une forme ronde à une structure de type vermiforme, une étroite bande de l'ectoderme se décolle de la ligne médiane dorsale pour devenir la plaque neurale, précurseur du système nerveux. La plaque neurale se creuse, s'invagine de manière à former la gouttière neurale puis, les plis neuraux qui bordent la gouttière fusionnent pour fermer la gouttière qui devient le tube neural. Ce tube se subdivise ensuite en une partie antérieure renflée, la vésicule céphalique primitive, qui se segmente en trois vésicules qui deviendront le prosencéphale, le mésencéphale, et le rhombencéphale44. Le prosencéphale se divise ensuite en deux autres vésicules, le télencéphale et le diencéphale tandis que le rhombencéphale se divise en métencéphale et myélencéphale. Chacune de ses vésicules contient des zones prolifératives dans lesquelles neurones et cellules gliales sont formés. Ces deux types de cellules migrent ensuite, parfois sur de longues distances, vers leurs positions finales.
Une fois qu'ils sont en place, les neurones commencent à étendre leurs dendrites et leur axone autour d'eux46. L'axone doit généralement s'étendre sur une longue distance à partir du corps cellulaire du neurone et doit se connecter sur des cibles bien spécifiques, ce qui lui nécessite de croître d'une manière plus complexe. À l'extrémité de l'axone en développement se trouve une région parsemée de récepteurs chimiques, le cône de croissance. Ces récepteurs recherchent des signaux moléculaires dans l'environnement alentour qui guident la croissance de l'axone en attirant ou en repoussant le cône de croissance et dirigent ainsi l'étirement de l'axone dans une direction donnée. Le cône de croissance navigue ainsi à travers le cerveau jusqu'à ce qu'il atteigne sa région de destination, où d'autres signaux chimiques engendrent la formation de synapses. Des milliers de gènes interviennent pour générer ces signaux de guidage mais le réseau synaptique qui en émerge n'est déterminé qu'en partie par les gènes. Dans de nombreuses parties du cerveau, les axones connaissent d'abord une surcroissance proliférative qui est ensuite régulée par des mécanismes dépendants de l'activité neuronale47. Ce processus sophistiqué de sélection et d'ajustement graduel aboutit finalement à la forme adulte du réseau neuronal.
Chez les mammifères les neurones sont produites avant la naissance (principalement entre la 6e et la 18e semaine gestationnelle chez l'humain). Le cerveau du nouveau-né contient donc substantiellement plus de neurones que celui de l'adulte car au cours du développement puis encore pendant le vieillissement, un grand nombre de ces cellules vont être détruites. La disparition des cellules nerveuses corresponda un phénomène nécessaire de sélection/stabilisation dans les réseaux de neurones au cours de la mise en place de circuits cérébraux.
Cependant quelques zones continuent de générer de nouveaux neurones tout au long de la vie, telles que le bulbe olfactif ou le gyrus dentatus de l'hippocampe. En dehors de ces exceptions, le nombre de neurones présents à la naissance est définitif, contrairement aux cellules gliales qui sont renouvelées tout au long de la vie, à la manière de la plupart des cellules de l'organisme. Bien que le nombre de neurones évolue peu après la naissance, les connexions axonales continuent de se développer et de s'organiser pendant encore un long moment. Chez l'Homme ce processus n'est pas terminé avant l'adolescence et il continue de se poursuivre avec l'acquisition de nouveaux apprentissages.
De nombreuses questions restent en suspens concernant ce qui relève de l'inné et de l'acquis à propos de l'esprit, de l'intelligence et de la personnalité48. Bien que de nombreux points restent à éclaircir, la neuroscience a montré que deux facteurs sont essentiels. D'un côté, les gènes déterminent la forme générale du cerveau, et la manière dont le cerveau répond à l'expérience. D'un autre côté, l'expérience est nécessaire pour affiner la matrice de connexions synaptiques. À bien des égards, la qualité et la quantité d'expériences joue un rôle49. L’enrichissement environnemental montre que le cerveau d'un animal placé dans un environnement plus riche et stimulant a un nombre plus important de synapses que celui d'un animal dans un milieu plus pauvre50.
Fonctions
La principale fonction du cerveau est de contrôler les actions de l'organisme à partir des informations sensorielles qui lui parviennent51. Les signaux sensoriels peuvent stimuler une réponse immédiate, moduler un schéma d'activité en cours, ou être emmagasinés pour un besoin futur. Ainsi, par le rôle central qu'il exerce dans la captation des stimuli externes, le cerveau occupe le rôle central dans la création de réponses à l'environnement. Le cerveau a aussi un rôle dans la régulation hormonale.
Le cerveau des vertébrés reçoit des signaux par les nerfs afférents de la part des différentes régions de l'organisme. Le cerveau interprète ces signaux et en tire une réponse fondée sur l'intégration des signaux électriques reçus, puis la transmet. Ce jeu de réception, d'intégration, et d'émission de signaux représente la fonction majeure du cerveau, qui explique à la fois les sensations, le mouvement, la mémoire et, on le suppose, la conscience.
Pour mener à bien sa complexe tâche, le cerveau est organisé en sous-systèmes fonctionnels c'est-à-dire que certaines régions cérébrales traitent plus spécifiquement certains aspects de l'information. Cette division fonctionnelle n'est pas stricte et ces sous-systèmes peuvent être catégorisés de plusieurs façons : anatomiquement, chimiquement ou fonctionnellement. Une de ces catégorisations repose sur les neurotransmetteurs chimiques utilisés par les neurones pour communiquer. Une autre se base sur la manière dont chaque zone du cerveau contribue au traitement de l'information : les zones sensorielles amènent l'information au cerveau ; les signaux moteurs envoient l'information du cerveau jusqu'aux muscles et aux glandes ; les systèmes excitateurs modulent l'activité du cerveau en fonction du moment de la journée et de divers facteurs.
Le cerveau utilise principalement le glucose comme substrat énergétique et une perte de conscience peut survenir s'il en manque. La consommation énergétique du cerveau n'est pas particulièrement variable, mais les régions actives du cortex consomment plus d'énergie que les inactives.
Systèmes de neurotransmissions
Selon le principe de Dale, chaque neurone du cerveau libère constamment le même neurotransmetteur chimique, ou la même combinaison de neurotransmetteurs, pour toutes les connexions synaptiques qu'il entretient avec d'autres neurones52. Un neurone peut donc être caractérisé en fonction des neurotransmetteurs qu'il libère bien qu'il existe quelques exceptions à ce principe. Les deux neurotransmetteurs les plus fréquents sont le glutamate, qui correspond généralement à un signal excitatoire, et l'acide γ-aminobutyrique (GABA), généralement inhibitoire. Les neurones utilisant ces deux neurotransmetteurs se retrouvent dans presque toutes les régions du cerveau et forment un large pourcentage des synapses du cerveau53.
Les autres neurotransmetteurs, comme la sérotonine ou la noradrénaline, proviennent de neurones localisés dans des zones particulières du cerveau. D'autres neurotransmetteurs, comme l'acétylcholine ou la dopamine, proviennent de plusieurs endroits du cerveau, mais ne sont pas distribués de façon aussi ubiquitaire que le glutamate et le GABA. La grande majorité des drogues psychotropes agissent en altérant les systèmes de neurotransmetteurs qui ne sont pas directement impliqués dans les transmissions glutamatergiques ou GABAergiques54.
Systèmes sensoriels
Une fonction importante du cerveau est de traiter l'information reçue par les récepteurs sensoriels55. Contrairement aux idées reçues, les sens que peut capter le cerveau ne sont pas limitées à cinq. Outre la vue, l'ouïe, le toucher, l'odorat, et le goût, le cerveau peut recevoir d'autres informations sensorielles comme la température, l'équilibre, la position des membres, ou la composition chimique du sang. Toutes ces variables sont détectées par des récepteurs spécialisés qui transmettent les signaux vers le cerveau. Certaines espèces peuvent détecter des sens supplémentaires, comme la vision infrarouge des serpents, ou utiliser les sens « standards » de manière non conventionnelle, comme l'écholocation du système auditif des chauves-souris.
Chaque système sensoriel possède ses propres cellules sensorielles réceptrices. Ces cellules sont des neurones mais, contrairement à la majorité des neurones, ceux-ci ne sont pas contrôlés par les signaux synaptiques d'autres neurones. Au lieu de cela, ces cellules sensorielles possèdent des récepteurs membranaires qui sont stimulées par un facteur physique spécifique comme la lumière, la température, ou la pression. Les signaux de ces cellules sensorielles réceptrices parviennent jusqu'à la moelle épinière ou le cerveau par les nerfs afférents.
Pour la plupart des sens, il y a un noyau sensitif principal dans le tronc cérébral, ou un ensemble de noyaux, qui reçoit et réunit les signaux des cellules sensorielles réceptrices. Dans de nombreux cas, des zones secondaires sous-corticales se chargent d'extraire et de trier l'information. Chaque système sensoriel a également une région du thalamus qui lui est dédié et qui relaie l'information au cortex.
Pour chaque système sensoriel, une zone corticale primaire reçoit directement les signaux en provenance du relai thalamique. Habituellement, un groupe spécifique de zones corticales supérieures analyse également le signal sensoriel. Enfin, des zones multimodales du cortex combinent les signaux en provenance de différents systèmes sensoriels. À ce niveau, les signaux qui atteignent ces régions du cerveau sont considérés comme des signaux intégrés plutôt que comme des signaux strictement sensoriels56.
Toutes ces étapes ont leurs exceptions. Ainsi, pour le toucher, les signaux sensoriels sont principalement reçus au niveau de la moelle épinière, au niveau de neurones qui projettent ensuite l'information au tronc cérébral57. Pour l'odorat, il n'y a pas de relai dans le thalamus, le signal est transmis directement de la zone primaire, le bulbe olfactif, vers le cortex58.
Systèmes moteurs
Les systèmes moteurs sont les zones du cerveau responsables directement ou indirectement des mouvements du corps, en agissant sur les muscles. À l'exception des muscles contrôlant les yeux, tous les muscles striés de l'organisme sont directement innervés par des neurones moteurs de la moelle épinière. Ils sont donc le dernier maillon de la chaîne du système psycho-moteur59. Les neurones moteurs spinaux sont contrôlés à la fois par des circuits neuronaux propres à la moelle épinière, et par des influx efférents du cerveau. Les circuits spinaux intrinsèques hébergent plusieurs réactions réflexes, ainsi que certains schémas de mouvements comme les mouvements rythmiques tels que la marche ou la nage60. Les connexions efférentes du cerveau permettent quant à elles, des contrôles plus sophistiqués.
Un certain nombre de zones du cerveau sont connectées directement à la moelle épinière61. Au niveau le plus inférieur se trouve les zones moteurs situées dans le bulbe rachidien et le pont. Au-dessus se situent les zones du mésencéphale, comme le noyau rouge, qui sont responsables de la coordination des mouvements. À un niveau supérieur se trouve le cortex moteur primaire, une bande de tissu cérébral localisé à la lisière postérieure du lobe frontal. Le cortex moteur primaire transmet ses commandes motrices aux zones moteurs sous-corticales, mais également directement à la moelle épinière par le biais du faisceau pyramidal. Les influx nerveux de ce faisceau cortico-spinal transmettent les mouvements fins volontaires. D'autres zones moteurs du cerveau ne sont pas directement reliées à la moelle épinière, mais agissent sur les zones moteurs primaires corticales ou sous-corticales. Quelques une de ces zones secondaires les plus importantes sont le cortex prémoteur, impliqués dans la coordination des mouvements de différentes parties du corps, les ganglions de la base, dont la fonction principale semble être la sélection de l'action, et le cervelet, qui module et optimise les informations pour rendre les mouvements plus précis.
Le cerveau et la moelle épinière contiennent également un réseau neuronal qui contrôle le système nerveux autonome, la partie du système nerveux responsable des fonctions automatiques. Non soumis au contrôle volontaire, le système nerveux autonome contrôle notamment la régulation hormonale et l'activité des muscles lisses et du muscle cardiaque. Le système nerveux autonome agit à différents niveaux comme le rythme cardiaque, la digestion, la respiration, la salivation, la miction, la sueur ou l'excitation sexuelle.
Systèmes d'éveil
Un des aspects les plus visibles du comportement animal est le cycle journalier veille-sommeil-rêve. L'éveil et l'attention sont aussi modulés à une échelle de temps plus fine, par un réseau de zones cérébrales62.
Un composant clé du système d'éveil est le noyau suprachiasmatique, petite région de l'hypothalamus localisée directement au-dessus du point de croisement des nerfs optiques63. Le noyau suprachiasmatique renferme l'horloge biologique centrale de l'organisme. Les neurones de ce noyau montrent un niveau d'activité qui augmente ou diminue sur une période d'environ 24 heures, le rythme circadien : cette activité fluctuante est dirigée par des changements rythmiques exprimés par un groupe de gènes horlogers. Le noyau suprachiasmatique reçoit généralement des signaux en provenance des nerfs optiques qui permettent de calibrer l'horloge biologique à partir des cycles jour-nuit.
Le noyau suprachiasmatique se projette dans un ensembles de zones cérébrales, situées au niveau de l'hypothalamus, du tronc cérébral, et du mésencéphale, qui sont impliqués dans la mise en œuvre des cycles jour-nuit. Un composant important du système est la formation réticulée, un groupe d'amas neuronaux s'étendant dans le tronc cérébral62. Les neurones réticulés envoient des signaux vers le thalamus, qui répond en envoyant des signaux à différentes régions du cortex qui régule le niveau d'activité.
Le sommeil implique de profondes modifications dans l'activité cérébrale64. Le cerveau ne s'éteint pas pendant le sommeil, l'activité cérébrale se poursuit mais est modifiée. En fait, il existe deux types de sommeil : le sommeil paradoxal (avec rêves) et le sommeil non paradoxal (généralement sans rêves). Ces deux sommeils se répètent selon un schéma légèrement différent à chaque sommeil. Trois grands types de schéma d'activité cérébrale peuvent être distingués : sommeil paradoxal, sommeil léger, et sommeil profond. Pendant le sommeil profond, l'activité du cortex prend la forme de larges ondes synchronisées tandis que ces ondes sont désynchronisées pendant l'état de rêve. Les niveaux de noradrénaline et de sérotonine tombent au cours du sommeil profond, et approchent du niveau zéro pendant le sommeil paradoxal, tandis que les niveaux d'acétylcholine présentent un schéma inverse.
Le cerveau et l'esprit
La compréhension de la relation entre le cerveau et l'esprit est un problème aussi bien scientifique que philosophique65. La relation forte entre la matière cérébrale physique et l'esprit est aisément mise en évidence par l'impact que les altérations physiques du cerveau ont sur l'esprit, comme le traumatisme crânien ou l'usage de psychotrope66.
Le problème corps-esprit est l'un des débat centraux de l'histoire de la philosophie et consiste à considérer la manière dont le cerveau et l'esprit sont reliés67. Trois grands courants de pensée existent concernant cette question : dualisme, matérialisme, et idéalisme. Le dualisme postule que l'esprit existe indépendamment du cerveau ; le matérialisme postule, quant à lui, que le phénomène mental est identique au phénomène neuronal ; et l'idéalisme postule que seul le phénomène mental existe68,69.
Outre ces questions philosophiques, la relation entre l'esprit et le cerveau soulève un grand nombre de questions scientifiques, comme la relation entre l'activité mentale et l'activité cérébrale, le mécanisme d'action des drogues sur la cognition, ou encore la corrélation entre neurones et conscience.
Historiquement, un grand nombre de philosophes considéraient inconcevable que la cognition puisse être mise en place par une substance physique comme le tissu cérébral70. Des philosophes comme Patricia Churchland ont postulé que l'interaction entre la drogue et l'esprit est un indicateur de la relation intime entre le cerveau et l'esprit mais que les deux entités sont distinctes71. René Descartes trouvait qu'il était possible d'expliquer les réflexes et d'autres comportements simples en termes mécaniques, mais il ne croyait pas que la pensée complexe, notamment le langage, puisse être expliqué par le cerveau physique uniquement72.
Antonio Damasio, dans son livre, L'erreur de Descartes, montre que le corps et l'esprit fonctionnent de manière indissociable et il explique que le raisonnement ne peut pas se faire sans les émotions73.
Recherche scientifique
Le domaine des neurosciences englobe toutes les approches cherchant à comprendre le fonctionnement du cerveau et du reste du système nerveux74. La psychologie cherche à comprendre l'esprit et le comportement. La neurologie est la discipline médicale qui diagnostique et traite les pathologies liées au système nerveux. Le cerveau est également l'organe le plus étudié en psychiatrie, une branche de la médecine qui étudie et traite les troubles mentaux75. Les sciences cognitives tentent de lier la neuroscience et la psychologie avec d'autres domaines comme l'informatique et la philosophie.
La plus ancienne méthode d'étude du cerveau est l'anatomie. Au milieu du XXe siècle, les progrès des neurosciences proviennent de l'amélioration des techniques de microscopie et de coloration76. Les neuroanatomistes étudient la structure du cerveau aussi bien à grande échelle qu'à l'échelle microscopique. Parmi d'autres outils, ils emploient une large gamme de colorants qui permettent de révéler la structure neurale, les réactions chimiques, et la connectivité. Le développement plus récent de techniques d'immunocoloration a permis de colorer les neurones qui exprime spécifiquement un groupe de gènes. Également, la neuroanatomie fonctionnelle utilise les techniques d'imagerie médicale pour corréler les variations dans la structure du cerveau avec les changements de cognition ou de comportement.
Les neurophysiologistes étudient les propriétés chimiques, pharmacologiques et électriques du cerveau. Leur principaux outils sont les drogues et les dispositifs d'enregistrement. Des milliers de drogues expérimentalement développées affectent le système nerveux, plusieurs le font de manière très spécifique. L'enregistrement de l'activité cérébrale peut se faire par l'utilisation d'électrodes, soit collées au crâne comme dans le cas d'électro-encéphalographie, soit implantées à l'intérieur du cerveau pour des enregistrements extracellulaires, qui peuvent détecter les potentiels d'action générés par des neurones individuels. Comme le cerveau ne contient pas de nocicepteurs, il est possible d'utiliser ces techniques sur un animal éveillé sans causer de douleur. Il est aussi possible d'étudier l'activité cérébrale par un examen non invasif en utilisant des techniques d'imagerie fonctionnelle comme l'IRM. Ainsi la tomographie à émission de positons met en évidence qu'en l'absence de toute focalisation particulière de l'attention, l'activité du cerveau (activité intrinsèque du réseau du mode par défaut, nommée « énergie sombre du cerveau » par analogie à l'énergie sombre du cosmos77 et qui consiste en des vagues d'ondes électriques lentes) correspond à une dépense de 60 à 80% de toute l'énergie consommée par le cerveau, soit une énergie 20 fois supérieure à celle consommée par le cerveau lorsqu'il réagit consciemment, la réalisation d'une tâche particulière (activite consciente moins fréquente que l'activité inconsciente) exige une énergie qui n'excède pas 5% de celle consommée par l'activité de fond78.
Une autre approche est d'examiner les conséquences de l'endommagement de zones spécifiques du cerveau. Bien que protégé par le crâne et les méninges, et isolé du flux sanguin par la Barrière hémato-encéphalique, le cerveau est tout de même vulnérable à de nombreuses maladies et à différents types de dégâts. Chez les humains, les effets des dégâts cérébraux sont une source importante d'informations sur la fonction cérébrale79. Comme il n'y a pas la capacité de contrôler expérimentalement la nature de ces dégâts, cette information est néanmoins souvent difficile à interpréter. Chez les animaux, les rats étant les plus fréquents sujets d'étude, il est possible d'utiliser des électrodes ou d'injecter localement des produits chimiques pour produire des types de dégâts bien précis et observer ensuite leurs conséquences sur le comportement.
Les neurosciences computationnelles regroupent deux approches : l'utilisation d'ordinateurs pour comprendre le cerveau et l'étude de la façon dont le cerveau réalise la computation80. D'un côté, il est possible de coder un programme informatique pour permettre de simuler le fonctionnement d'un groupe de neurones en utilisant des systèmes d'équations décrivant l'activité électrochimique ; ces simulations sont appelées « réseaux de neurones biologiquement réalistes ». D'un autre côté, il est possible d'étudier les algorithmes de computation neurale par la simulation ou l'analyse mathématique d'« unités » simplifiées ayant plusieurs des caractéristiques des neurones mais en faisant abstraction de la plupart de leur complexité biologique. Les fonctions computationnelles du cerveau sont étudiés à la fois par les neuroscientifiques et les informaticiens.
Les dernières années ont vu les premières applications des techniques d'ingénierie génétique pour l'étude du cerveau81. Les sujets d'études les plus fréquents sont les souris, car c'est sur cette espèce que les outils techniques à disposition sont le plus au point. Il est désormais possible d'inactiver ou de muter une grande variété de gènes, et ensuite examiner les effets sur la fonction cérébrale. Des approches plus sophistiquées sont également utilisées, comme la recombinaison Cre-Lox qui permet d'activer ou d'inactiver les gènes dans des zones spécifiques du cerveau à des moments spécifiques.
Des équipes américaines, bénéficiant d'un financement fédéral de 28 millions d'euros, utilisent des scanners dernier cri pour créer une banque de « connectomes ». Ces cartes des circuits cérébraux promettent de révéler comment les organes réagissent au vieillissement, à l'apprentissage et à d'autres événements. Les données du Human Connectome Project laisseraient présager des avancées sur les traitements de l'autisme et de la schizophrénie.
Histoire
Historiquement, les avis se sont souvent opposés pour savoir qui du cerveau ou du cœur était le siège de l'âme. Dans un sens, il était impossible de nier que la conscience semble comme localisée dans la tête, qu'un coup à la tête provoque bien plus facilement l'inconscience qu'un coup sur le torse, et que secouer la tête donne des vertiges. Dans un autre sens, le cerveau soumis à un examen superficiel semble inerte, tandis que le cœur bat constamment. L'arrêt des battements cardiaques provoque la mort, tandis que les émotions induisent des changements du rythme cardiaque et que le chagrin produit souvent une sensation de douleur au niveau du cœur (« cœur brisé »). Pour Aristote, le siège de l'âme était le cœur et le cerveau, organe de refroidissement, servait simplement à assurer la circulation du sang (théorie philosophique et médicale du cardiocentrisme). Démocrite divise l'âme en trois parties : l'intellect dans la tête, l'émotion dans le cœur, le désir autour du foie82. Hippocrate était sûr que l'âme se trouvait dans le cerveau (théorie philosophique et médicale du cérébrocentrisme). Pour Hérophile, le cerveau était le centre de l’intelligence83.

Dessin de Santiago Ramón y Cajal de neurones de pigeon observés grâce à la coloration de Golgi.
Galien a aussi défendu l'importance du cerveau, et élaboré des théories assez avancées sur sa manière de fonctionner. Même après la reconnaissance de la primauté du cerveau par la société savante, l'idée du cœur comme siège de l'intelligence a continué de survivre dans les idiomes populaires84. Galien a entrepris un long travail pour décrire les relations anatomiques entre le cerveau, les nerfs, et les muscles, démontrant que tous les muscles du corps sont connectés au cerveau par un réseau de nerfs. Il postulait que les nerfs activent les muscles mécaniquement, en transportant une substance mystérieuse qu'il appelle le pneuma psychique (ou « esprit animal »). Ses idées connaissent une relative renommée pendant le Moyen-Âge, mais ne progressent pas jusqu'à la Renaissance. À la Renaissance, des études anatomiques détaillées résument et combinent les idées de Galien avec celles de Descartes et ses successeurs. Comme Galien, Descartes pensait le système nerveux en termes hydrauliques. Il est convaincu que les plus hautes fonctions cognitives, le langage notamment, sont effectuées par une « chose pensante » non-physique (res cogitans), mais que la majorité des comportements humains et animaux peuvent s'expliquer mécaniquement. Le premier grand pas vers la médecine moderne provient des recherches de Luigi Galvani, qui découvre qu'une décharge d'électricité statique appliquée sur le nerf d'une grenouille morte provoque la contraction de sa jambe.
Chaque grande avancée dans la compréhension du cerveau a suivi plus ou moins directement le développement d'une nouvelle méthode d'analyse. Au XVIIe siècle, Thomas Willis, père des neurosciences modernes prend le contrepied de la théorie dualiste de Descartes, privilégiant l'approche matérialiste grâce à sa description du fonctionnement des nerfs. Au XVIIIe siècle, Vincenzo Malacarne (it), fondateur de l'anatomie topographique et de la neuropsychologie, pose les principes de l'existence de signes cliniques de ses patients reliés à l’observation post-mortem de lésions de leur cerveau. Paul Broca démontre cette corrélation anatomo-clinique85. Au XIXe siècle, Franz Gall développe grâce à des techniques organologiques, une théorie localisationiste selon laquelle les fonctions cognitives sont au niveau des circonvolutions du cortex cérébral86. Les chirurgiens Charles Scott Sherrington et C.S. Roy observent le cerveau en salle d'opération et remarquent qu'une zone du cortex rougit lorsque le patient bouge une partie précise de son corps, montrant la consommation d'oxygène cérébrale.
Jusqu'au début du XXe siècle, les principales avancées ont découlé de la découverte de nouveaux colorants histologiques87. Sans coloration, le tissu cérébral apparait au microscope comme un impénétrable enchevêtrement de fibres protoplasmiques dans lequel il était impossible de déterminer une quelconque structure. L'une des colorations les plus importantes est la coloration de Golgi qui permet de colorer seulement une petite fraction de neurones. La coloration des neurones se fait apparemment de façon aléatoire mais colore les neurones dans leur entier, aussi bien le corps cellulaire que les dendrites et l'axone. Grâce notamment aux travaux de Camillo Golgi et de Santiago Ramón y Cajal, la nouvelle coloration a pu révéler des centaines de types distincts de neurones, chacun avec sa propre structure dendritique et son propre schéma de connectivité.
Au XXe siècle, les progrès en électronique ont ouvert la voie à la recherche des propriétés électriques des cellules nerveuses, notamment aux travaux comme ceux de Louis Sokoloff sur l'autoradiographie (le débit sanguin cérébral par la méthode des traceurs radioactifs permet de réaliser une cartographie fonctionnelle88), d'Alan Hodgkin ou d'Andrew Huxley sur la biophysique du potentiel d'action, et les travaux sur l'électrochimie de la synapse comme ceux de Bernard Katz89. Les premières études utilisaient des préparations spéciales, comme avec l'étude de la réponse de fuite rapide du calmar qui fait intervenir un axone et des synapses géantes. L'amélioration constante des électrodes et de l'électronique a permis d'affiner le niveau de résolution des recherches. Ces études ont complété les planches anatomiques en concevant le cerveau comme une entité dynamique.
Aux États-Unis, les années 1990 sont déclarées « Décennie du cerveau » pour commémorer les avancées de la recherche sur le cerveau, et promouvoir les fonds pour les recherches futures90,91.
Notes et références
- ↑ a et b (en) G. M. Shepherd, Neurobiology, Oxford University Press, 1994, 3e éd., 760 p. (ISBN 9780195088434) [présentation en ligne [archive]], p. 3
- ↑ (en) G. Balavoine et A. Adoutte, « The Segmented Urbilateria: A Testable Scenario », dans Integr. Comp. Biol., vol. 43, no 1, 2003, p. 137-147 (ISSN 1540-7063 et 1557-7023) [texte intégral [archive], lien DOI [archive]]
- ↑ (en) A. Schmidt-Rhaesa, The evolution of organ systems, Oxford University Press, 2007, 385 p. (ISBN 9780198566694) [présentation en ligne [archive]], p. 110
- ↑ a et b (en) A. B. Butler, « Chordate evolution and the origin of craniates: An old brain in a new head », dans Anat. Rec., vol. 261, no 3, 15 juin 2000, p. 111-125 (ISSN 1932-8494) [lien DOI [archive]]
- ↑ (en) E. R. Kandel, In search of memory: the emergence of a new science of mind, W. W. Norton & Co., 2007, 510 p. (ISBN 9780393329377) [présentation en ligne [archive]]
- ↑ Flybrain: An online atlas and database of the drosophila nervous system [archive], Département d'entomologie de l'université du Colorado. Consulté le 29 octobre 2010
- ↑ (en) R. J. Konopka et S. Benzer, « Clock Mutants of Drosophila melanogaster », dans Proc. Natl. Acad. Sci. USA, vol. 68, no 9, 1er septembre 1971, p. 2112-2116 (ISSN 0027-8424) [texte intégral [archive]]
- ↑ (en) H.-S. Shin, T. A. Bargiello, B. T. Clark, F. R. Jackson et M. W. Young, « An unusual coding sequence from a Drosophila clock gene is conserved in vertebrates », dans Nature, vol. 317, no 6036, 3 octobre 1985, p. 445-448 (ISSN 0028-0836) [lien DOI [archive]]
- ↑ WormBook: The online review of C. Elegans biology [archive]. Consulté le 30 octobre 2010
- ↑ (en) O. Hobert, « Specification of the nervous system », dans WormBook, The C. elegans Research Community, 8 août 2005, p. 1-19 [texte intégral [archive], lien DOI [archive] (pages consultées le 26 décembre 2010)]
- ↑ (en) Sydney Brenner [archive], 1987. Consulté le 26 décembre 2010
- ↑ (en) J. G. White, E. Southgate, J. N. Thomson et S. Brenner, « The Structure of the Nervous System of the Nematode Caenorhabditis elegans », dans Phil. Trans. R. Soc. B, vol. 314, no 1165, 12 novembre 1986, p. 1-340 (ISSN 1471-2970) [lien DOI [archive]]
- ↑ (en) D.-G. Shu, S. Conway Morris, J. Han, Z.-F. Zhang, K. Yasui, P. Janvier, L. Chen, X.-L. Zhang, J.-N Liu, Y. Li et H.-Q Liu, « Head and backbone of the Early Cambrian vertebrate Haikouichthys », dans Nature, vol. 421, no 6922, 30 janvier 2003, p. 526-529 (ISSN 0028-0836) [lien DOI [archive]]
- ↑ R. Eckert et D. Randall (trad. F. Math), Physiologie animale : Mécanismes et adaptations, De Boeck Université, 1999, 4e éd., 822 p. (ISBN 9782744500534) [lire en ligne [archive] (page consultée le 1er janvier 2011)] [présentation en ligne [archive]], p. 415
- ↑ (en) G. F. Striedter, Principles of brain evolution, Sinauer Associates, 2005, 436 p. (ISBN 9780878938209) [présentation en ligne [archive]]
- ↑ (en) E. R. Kandel, J. H. Schwartz et T. M. Jessell, Principles of neural science, McGraw-Hill, 2000, 4e éd., 1414 p. (ISBN 9780838577011) [présentation en ligne [archive]], chap. 17
- ↑ (en) A. Parent et M. B. Carpenter, Carpenter's human neuroanatomy, Williams & Wilkins, 1996, 9e éd., 1011 p. (ISBN 9780683067521) [présentation en ligne [archive]], chap. 1
- ↑ Kandel 2000, p. 1019
- ↑ a et b Jean-Didier Vincent Pierre-Marie Liedo, Le cerveau sur mesure, Odile Jacob, 2012, 290 p. (ISBN 2738127096)
- ↑ (en) E. Armstrong, « Relative Brain Size and Metabolism in Mammals », dans Science, vol. 220, no 4603, 17 juin 1983, p. 1302-1304 (ISSN 0036-8075) [lien DOI [archive]]
- ↑ a et b (en) B. L. Finlay, R. B. Darlington et N. Nicastro, « Developmental structure in brain evolution », dans Behav. Brain Sci., vol. 24, no 2, 2001, p. 263-278 [lien DOI [archive]]
- ↑ Kandel 2000, ch. 17
- ↑ (en) R. G. Northcutt, « Forebrain evolution in bony fishes », dans Brain Res. Bull., vol. 75, no 2-4, 18 mars 2008, p. 191-205 (ISSN 0361-9230) [lien DOI [archive]]
- ↑ (en) A. Reiner, K. Yamamoto et H. J. Karten, « Organization and evolution of the avian forebrain », dans Anat. Rec., vol. 287A, no 1, novembre 2005, p. 1080–1102 (ISSN 1932-8494) [lien PMID [archive], lien DOI [archive]]
- ↑ Kandel 2000, ch. 44-45
- ↑ (en) D. F. Swaab, The human hypothalamus : Handbook of clinical neurology. Neuropathology of the human hypothalamus and adjacent brain structure, vol. 2, Elsevier Health Sciences, 2004, 597 p. (ISBN 9780444514905) [présentation en ligne [archive]]
- ↑ (en) E. G. Jones, The thalamus, Plenum Press, 1985, 935 p. (ISBN 9780306418563) [présentation en ligne [archive]]
- ↑ Kandel 2000, ch. 42
- ↑ (en) K. Saitoh, A. Ménard et S. Grillner, « Tectal control of locomotion, steering, and eye movements in lamprey », dans J. Neurophysiol., vol. 97, no 4, avril 2007, p. 3093-3108 (ISSN 0022-3077) [texte intégral [archive], lien PMID [archive], lien DOI [archive]]
- ↑ (en) L. Puelles, « Thoughts on the development, structure and evolution of the mammalian and avian telencephalic pallium », dans Phil. Trans. R. Soc. B, vol. 356, no 1414, 29 octobre 2001, p. 1583-1598 (ISSN 0962-8436) [texte intégral [archive], lien PMID [archive], lien DOI [archive]]
- ↑ (en) C. Salas, C. Broglio et F. Rodríguez, « Evolution of Forebrain and Spatial Cognition in Vertebrates: Conservation across Diversity », dans Brain Behav. Evol., vol. 62, no 2, 2003, p. 72-82 (ISSN 0006-8977 et 1421-9743) [lien PMID [archive], lien DOI [archive]]
- ↑ S. Grillner, J. Hellgren, A. Ménard, K. Saitoh et M. A. Wikström, « Mechanisms for selection of basic motor programs – roles for the striatum and pallidum », dans Trends Neurosci., vol. 28, no 7, juillet 2005, p. 364-370 (ISSN 0166-2236) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) R. G. Northcutt, « Evolution of the Telencephalon in Nonmammals », dans Ann. Rev. Neurosci., vol. 4, mars 1981, p. 301-350 (ISSN 0147-006X) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) R. A. Barton et P. H. Harvey, « Mosaic evolution of brain structure in mammals », dans Nature, vol. 405, no 6790, 29 juin 2000, p. 1055-1058 (ISSN 0028-0836 et 1476-4687) [lien PMID [archive], lien DOI [archive]]
- ↑ a et b (en) F. Aboitiz, D. Morales et J. Montiel, « The evolutionary origin of the mammalian isocortex: Towards an integrated developmental and functional approach », dans Behav. Brain Sci., vol. 26, no 5, 2003, p. 535-552 (ISSN 0140-525X) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) W. H. Calvin, How brains think : Evolving intelligence, then and now, Basic Books, 1997, 192 p. (ISBN 9780465072781 et 046507278X) [présentation en ligne [archive]]
- ↑ (en) M. I. Sereno, A. M. Dale, J. B. Reppas, K. K. Kwong, J. W. Belliveau, T. J. Brady, B. R. Rosen et R. B. Tootell, « Borders of multiple visual areas in humans revealed by functional magnetic resonance imaging », dans Science, vol. 268, no 5212, 12 mai 1995, p. 889-893 (ISSN 0036-8075 et 1095-9203) [texte intégral [archive], lien PMID [archive], lien DOI [archive]] :
« Over half of the neocortex in non-human primates is occupied by visual areas. At least 25 visual areas beyond the primary visual cortex (V1) have been identified with a combination of microelectrode mapping, tracer injections, histological stains, and functional studies »
- ↑ Kandel 2000, p. 20
- ↑ (en) F. A. C. Azevedo, L. R. B. Carvalho, L. T. Grinberg, J. M. Farfel, R. E. L. Ferretti, R. E. P. Leite, W. J. Filho, R. Lent et s. Herculano-Houzel, « Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain », dans J. Comp. Neurol., vol. 513, no 5, 10 avril 2009, p. 532–541 (ISSN 1096-9861) [lien PMID [archive], lien DOI [archive]]
- ↑ Kandel 2000, p. 21
- ↑ Kandel 2000, ch. 10
- ↑ Kandel 2000, ch. 2
- ↑ (en) L. Marner, J. R. Nyengaard, Y. Tang et B. Pakkenberg, « Marked loss of myelinated nerve fibers in the human brain with age », dans J. Comp. Neurol., vol. 462, no 2, 21 juillet 2003, p. 144-152 (ISSN 1096-9861) [lien PMID [archive], lien DOI [archive]]
- ↑ a, b et c (en) D. Purves et J. W. Lichtman, Principles of neural development, Sinauer Associates, 1985, 433 p. (ISBN 9780878937448) [présentation en ligne [archive]], chap. 1
- ↑ Purves 1985, ch. 4
- ↑ Purves 1985, ch. 5 et 7
- ↑ Purves 1985, ch. 12
- ↑ (en) M. Ridley, The agile gene : How nature turns on nurture, Forth Estate, 2004, 352 p. (ISBN 9780060006792) [présentation en ligne [archive]]
- ↑ (en) T. N. Wiesel, « Postnatal development of the visual cortex and the influence of environment », dans Nature, vol. 299, no 5884, 14 octobre 1982, p. 583-591 (ISSN 0028-0836 et 1476-4687) [texte intégral [archive], lien PMID [archive], lien DOI [archive]]
- ↑ (en) H. van Praag, G. Kempermann et F. H. Gage, « Neural consequences of enviromental enrichment », dans Nat. Rev. Neurosci., vol. 1, no 3, décembre 2000, p. 191-198 (ISSN 1471-003X et 1471-0048) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) T. J. Carew, Behavioral neurobiology : The cellular organization of natural behavior, Sinauer Associates, 2000, 435 p. (ISBN 9780878930920) [présentation en ligne [archive]], chap. 1
- ↑ Kandel 2000, ch. 15
- ↑ (en) P. L. McGeer et E. G. McGeer, Basic neurochemistry : Molecular, cellular and medical aspects, New York, Raven Press, 1989, 4e éd., 984 p. (ISBN 9780881673432) [présentation en ligne [archive]], chap. 15 (« Amino acid neurotransmitters »), p. 311-332
- ↑ (en) J. R. Cooper, F. E. Bloom et R. H. Roth, The biochemical basis of neuropharmacology, Oxford University Press, 2003, 8e éd., 405 p. (ISBN 9780195140088) [présentation en ligne [archive]]
- ↑ Kandel 2000, ch. 21
- ↑ Kandel 2000, ch. 21 et 30
- ↑ Kandel 2000, ch. 23
- ↑ Kandel 2000, ch. 32
- ↑ Kandel 2000, ch. 34
- ↑ Kandel 2000, ch. 36-37
- ↑ Kandel 2000, ch. 33
- ↑ a et b Kandel 2000, ch. 45
- ↑ (en) M. C. Antle et R. Silver, « Orchestrating time: arrangements of the brain circadian clock », dans Trends Neurosci., vol. 28, no 3, mars 2005, p. 145-151 (ISSN 0166-2236) [texte intégral [archive], lien PMID [archive], lien DOI [archive]]
- ↑ Kandel 2000, ch. 47
- ↑ (en) P. S Churchland, Neurophilosophy : Toward a unified science of the mind-brain, MIT Press, 1990, 5e éd., 546 p. (ISBN 9780262530859) [présentation en ligne [archive]]
- ↑ (en) C. Boake et L. Dillar, Rehabilitation for Traumatic Brain Injury, Oxford University Press, juin 2005, 385 p. (ISBN 9780195173550), chap. 1 (« History of Rehabilitation for Traumatic Brain Injury »)
- ↑ Churchland 1990, ch. 7
- ↑ (en) W. D. Hart et S. Guttenplan (dir.), A companion to the philosophy of mind, Wiley-Blackwell, 1996, 656 p. (ISBN 9780631199960) [lire en ligne [archive]], « Dualism », p. 265-267
- ↑ (en) A. R. Lacey, A dictionary of philosophy, Routledge, 1996, 3e éd. (1re éd. 1976), 386 p. (ISBN 9780415133326) [lire en ligne [archive]]
- ↑ Churchland 1990, ch. 6
- ↑ Churchland 1990, ch. 8
- ↑ (en) R. Descartes (trad. R. Stoothoff, D. Murdoch, J. Cottingham), The philosophical writings of Descartes, Cambridge University Press, 1991, 448 p. (ISBN 9780521288088)
- ↑ Damasio et Blanc 2006.
- ↑ Kandel 2000, ch. 1
- ↑ (en) H. A. Storrow, Outline of clinical psychiatry, Appleton-Century-Crofts, 1969 (ISBN 9780390850751)
- ↑ (en) M. F. Bear, B. W. Connors et M. A. Paradiso, Neuroscience : Exploring the brain, Lippincott Williams & Wilkins, 3e éd., 857 p. (ISBN 9780781760034) [lire en ligne [archive]], chap. 2 (« Neurons and glia »)
- ↑ (en) Marcus Raichle, « The Brain's Dark Energy », dans Science, vol. 314, 2009, p. 1249-1250
- ↑ Marcus Raichle (en), « Un cerveau jamais au repos », dans Pour la Science, no 393, 2010, p. 42-47
- ↑ (en) B. Kolb et I. Q. Whishaw, Fundamentals of human neuropsychology, Worth Publisherslieu=, 2008, 6e éd., 763 p. (ISBN 9780716795865) [présentation en ligne [archive]], chap. 1
- ↑ (en) P. Dayan et L. F. Abbott, Theoretical neuroscience : Computational and mathematical modeling of neural systems, MIT Press, 2005, 460 p. (ISBN 9780262541855) [présentation en ligne [archive]]
- ↑ (en) S. Tonegawa, K. Nakazawa et M. A. Wilson, « Genetic neuroscience of mammalian learning and memory », dans Phil. Trans. R. Soc. B, vol. 358, no 1432, 29 avril 2003, p. 787-795 (ISSN 1471-2970) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) S. Finger, Origins of neuroscience : A history of explorations into brain function, Oxford University Press, 2001, 480 p. (ISBN 9780195146943) [présentation en ligne [archive]], p. 14
- ↑ De la psychobiologie aux neurosciences cognitives [archive][PDF]
- ↑ (en) R. Hendrickson, The facts on file encyclopedia of word and phrase origins, New York, Facts on File, 2008, 4e éd., 948 p. (ISBN 9780816069668) [présentation en ligne [archive]]
- ↑ Modèles cognitifs de la mémoire [archive][PDF]
- ↑ Bernard Mazoyer, John W. Belliveau, « Les nouveaux progres de l'imagerie [archive] » sur larecherche.fr, 1986
- ↑ (en) F. E. Bloom et J. P. Swazey (dir.), The Neurosciences, paths of discovery, vol. 1973, MIT Press, 1975, 622 p. (ISBN 9780262230728) [présentation en ligne [archive]], p. 211
- ↑ (en) Louis Sokoloff, « The metabolism of the central nervous system in vivo », dans Neurophysiology, vol. 3, 1960, p. 1843–1864
- ↑ (en) M. Piccolino, « Fifty years of the Hodgkin-Huxley era », dans Trends Neurosci., vol. 25, no 11, 1er novembre 2002, p. 552-553 (ISSN 0166-2236) [lien PMID [archive], lien DOI [archive]]
- ↑ (en) George H. W. Bush, « Project on the Decade of the Brain [archive] » sur http://www.loc.gov/ [archive], juillet 1990. Consulté le 10 novembre 2010
- ↑ (en) E. G. Jones et L. M. Mendell, « Assessing the decade of the brain », dans Science, vol. 284, no 5415, 30 avril 1999, p. 739 (ISSN 0036-8075 et 1095-9203) [lien DOI [archive]]
Références bibliographiques
- Alonso-Nanclares L, Gonzalez-Soriano J, Rodriguez JR, DeFelipe J, « Gender differences in human cortical synaptic density », dans Proc Nat Acad Sci U.S.A., vol. 105, no 38, 2008, p. 14615–9 [lien PMID, lien DOI]
- (en) DD Clark, Basic Neurochemistry: Molecular, Cellular and Medical Aspects, Philadelphia, Lippincott, 1999 (ISBN 9780397518203), p. 637–70
- (en) JE Dowling, Neurons and Networks, Cambridge, Harvard University Press, 2001, 2e éd. (ISBN 978-0-674-00462-7) (LCCN 00065134) [lire en ligne]
- WJ Gehring, « New Perspectives on Eye Development and the Evolution of Eyes and Photoreceptors: The Evolution of Eyes and Brain. », dans J Heredity, vol. 96, no 3, 2005, p. 171–184 [texte intégral [Full text], lien PMID, lien DOI (pages consultées le 2008-04-26)]
- S Grillner, « Cellular bases of a vertebrate locomotor system-steering, intersegmental and segmental co-ordination and sensory control. », dans Brain Res Brain Res Rev, vol. 40, no 1-3, 2002, p. 92–106 [lien PMID, lien DOI]
- K Gurney, « Computational models of the basal ganglia: from robots to membranes. », dans Trends Neurosci, vol. 27, no 8, 2004, p. 453–9 [lien PMID, lien DOI]
- (en) Hippocrates, On the Sacred Disease, 400 B.C.E. [lire en ligne]
- (en) HJ Jerison, Evolution of the Brain and Intelligence, New York, Academic Press, 1973 (ISBN 978-0-12-385250-2) (LCCN 73002062)
- (en) N Kleitman, Sleep and Wakefulness, Chicago, The University of Chicago Press, Midway Reprints series, 1938, revised 1963, reprinted 1987 (ISBN 978-0-226-44073-6) (OCLC 18282650) (LCCN 63017845)
- M Nickel, « Dynamics and cellular movements in the locomotion of the sponge Tethya wilhelma. », dans Integr Comp Biol, vol. 42, 2002, p. 1285
- T Paus, « Maturation of white matter in the human brain: a review of magnetic resonance studies. », dans Brain Res Bull, vol. 54, no 3, 2001, p. 255–66 [lien PMID, lien DOI]
- Pelvig, P.; Pakkenberg, H.; Stark, K.; Pakkenberg, B. (Nov 2008). "Neocortical glial cell numbers in human brains". Neurobiology of aging 29 (11): 1754–1762. doi:10.1016/j.neurobiolaging.2007.04.013. ISSN 0197-4580. PMID 17544173
- M Raichle, « Appraising the brain's energy budget », dans Proc Nat Acad Sci U.S.A., vol. 99, no 16, 2002, p. 10237–9 [lien PMID, lien DOI]
- K Safi, « Bigger is not always better: when brains get smaller », dans Biol Lett, vol. 1, no 3, 2005, p. 283–6 [lien PMID, lien DOI]
- MV Savage, « The predominance of quarter-power scaling in biology », dans Functional Ecol, vol. 18, 2004, p. 257–82 [lien DOI]
- (en) J Searle, Mind, Language and Society, New York, Basic Books, 1999, 1re éd. (ISBN 978-0-465-04521-1) (LCCN 99187320) [lire en ligne]
- (en) JL van Hemmen, Sejnowski TJ, 23 Problems in Systems Neuroscience, Oxford, Oxford University Press, 2005 (ISBN 978-0-19-514822-0) [lire en ligne]
- (en) J von Neumann, Churchland PM, Churchland PS, The computer and the brain, New Haven, Yale Univ. Press, 2000, 2e éd., poche (ISBN 978-0-300-08473-3) (LCCN 00026937)
- (en) Willmer P, Invertebrate Relationships: Patterns in Animal Evolution, Cambridge, Cambridge University Press, 1990, 1re éd. (ISBN 978-0-521-33712-0) (LCCN 87032019) [lire en ligne]
- R Wong, « Retinal waves and visual system development. », dans Ann Rev Neurosci, vol. 22, 1999, p. 29–47 [lien PMID, lien DOI]
- (en) E Wyllie, Gupta A, Lachhwani DK, The Treatment of Epilepsy: Principles and Practice, Philadelphie, Lippincott Williams & Wilkins, 2005, 4e éd. (ISBN 978-0-7817-4995-4) (LCCN 2005022784) [lire en ligne]
Voir aussi
Sur les autres projets Wikimedia :
Bibliographie
- P. Buser et R. Lestienne, Cerveau, information, connaissance, CNRS Éditions, 2001, 227 p. (ISBN 9782271057730) [présentation en ligne]
- J.-P. Changeux, Du vrai, du beau, du bien : Une nouvelle approche neuronale, Odile Jacob, 2008, 544 p. (ISBN 9782738119049) [présentation en ligne]
- A. R. Damasio, L'autre moi-même : Les nouvelles cartes du cerveau, de la conscience et des émotions, Odile Jacob, 2010 (ISBN 9782738119407) [présentation en ligne]
- A. R. Damasio et M. Blanc, L'erreur de Descartes : La raison des émotions, Odile Jacob, 2006, 368 p. (ISBN 9782738117137) [présentation en ligne]
- J.-J. Feldmeyer, Le cerveau, Le Cavalier Bleu, coll. « Idées reçues » (no 131), 2007, 127 p. (ISBN 9782846701556) [présentation en ligne]
- O. Houdé, B. Mazoyer, N. Tzourio-Mazoyer et F. Crivello, Cerveau et psychologie : Introduction à l'imagerie cérébrale anatomique et fonctionnelle, Presses universitaires de France, 2002, 609 p. (ISBN 9782130509127) [présentation en ligne]
- M. Imbert, Traité du cerveau, Odile Jacob, 2006, 532 p. (ISBN 9782738116659) [présentation en ligne]
- J. E. LeDoux (trad. P. Kaldy), Le cerveau des émotions : Les mystérieux fondements de notre vie émotionnelle, Odile Jacob, 2005, 374 p. (ISBN 9782738116338) [présentation en ligne]
- M. Jeannerod, Le cerveau volontaire, Odile Jacob, 2009, 297 p. (ISBN 9782738123572) [présentation en ligne]
- R. Jouvent, Le cerveau magicien : De la réalité au plaisir psychique, Odile Jacob, 2009, 252 p. (ISBN 9782738118790) [présentation en ligne]
- B. Kolb et I. Q. Whishaw, Cerveau et comportement [« Neurosciences & cognition »], De Boeck Université, 2002, 672 p. (ISBN 9782744501371) [présentation en ligne]
- J.-D. Vincent, Voyage extraordinaire au centre du cerveau, Odile Jacob, 2007, 455 p. (ISBN 9782738119353) [présentation en ligne]
- J.-D. Vincent, Biologie des passions, Odile Jacob, 1999, 341 p. (ISBN 9782738107374) [présentation en ligne]
Articles connexes
Liens externes



CULTURE A NICE ... CULTURE A NICE ...
Chers Amis,
Vous êtes chaleureusement invités à une Fête organisée au Museaav
le vendredi 12 octobre à 18H30.
Nous vous présenterons:
- La nouvelle co-présidence représentée par Laurence Navalesi & Enrico Pastorini ainsi que la nouvelle équipe du MUSEAAV.
- Nos expositions actuelles du photographe-médium Wyl, les aquarelles de l’artiste allemand Udo Dietrich et la rétrospective de Bo Breguet, présentées dans nos locaux fraîchement repeints.
- Notre programmation toujours aussi variée, pour la fin de l’année 2012.
- Nos projets ambitieux pour l’année 2013.
Nous conclurons notre soirée par un verre de l’amitié.
L’EQUIPE DU MUSEAAV
LES PHOTOS DE LA SOIREE AVEC
AGENCY PRESS INTERNATIONAL DIACONESCO.TV
C. COPYRIGHTS ALL RIGHTS - NICE - 12.10.2012

















Photographe de Presse Gérard Diaconesco C. Copyrights Agency Press International DIACONESCO.TV
Publié le Conférences, Expo Temporaire, Performances |
Publié le Conférences, Expo Temporaire |
Publié le Conférences, Expo Temporaire, Performances |

Inauguration officielle samedi 20 octobre à 11H30 – Cocktail clôture vendredi 26 Octobre.
2012 est l’année de la langue russe en France.
L’organisation à Nice d’une semaine dédiée à ce thème est l’occasion pertinente d’attirer l’attention des pouvoirs publics et des habitants de notre région sur la nécessité de favoriser l’enseignement du russe (du primaire au supérieur) pour répondre aux besoins économiques et culturels du public français autant dans la valorisation de leur savoir pour leur profession, que pour la communication et l’accès à la culture. C’est aussi l’occasion pour les russophones résidant dans notre région de garder le lien avec leur culture originelle.
Pour notre région, cet événement permettra de faire le point sur la présence russe, d’attirer des villes voisines de nombreux visiteurs, russophones et francophones, et, au-delà de la Région, grâce à la présence d’exposants de la France entière, de développer des coopérations et des échanges pour l’édition, la traduction, la création de journaux ou de télévisions tournés résolument vers la modernité, sans oublier les racines profondes de nos deux cultures.
Expositions de Tatyana Danilyands : L’Âme russe- Photos et sculptures en verre
- Elana : Peintures
Ateliers marionnettes, chants et danses russes, court métrage et poésie, atelier de contes, initiation à la langue russe, littérature sur la présence russe à Nice
PROGRAMME
| Date |
Contenu |
Lieu |
|
4 octobre
|
|
|
|
19h00
|
Conférence. Les Académies des Saisons Musicales Russes à Nice. |
Eglise Anglicane de Nice |
|
20h00
|
Concert. Violons Virtuoses. Vladimir Ivanov (violon), Elena Chvertok (violon), Elena Keylina (orgue). Corelli, Haendel, Bach |
Eglise Anglicane de Nice11, rue de la Buffa |
|
19 octobre
|
|
|
|
21h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La pelisse » N.Gogol |
Théâtre de la Cité |
| 20 – 27 octobre Semaine de la langue et des lettres russes au MUSEEAV |
|
20 octobre
|
|
|
|
14h -18h
|
Expositions : Accueil du public, visite des stands, visite des expositions, rencontre avec les artistes exposants (Elana, Tatyana Danilyants…, rencontre interprètes traducteurs) |
MUSEAAV |
|
14h00 – 15h30
|
Rencontre avec Pavel Komarov (Zertsala) et les acteurs de nos diverses troupes |
MUSEAAV |
|
16h – 18h00
|
Tatyana Danilyants : « l’Ame russe », photos et cinéma. |
|
|
21h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La pelisse » N.Gogol |
Théâtre de la Cité |
|
21 octobre
|
|
|
|
10h -18h
|
Accueil du public, visite des stands, visite des expositions. rencontre avec les enseignants de langues russe et française étrangères, les libraires, les éditeurs ; tables rondes avec les écrivains sur les questions de littérature, de traduction, de la langue russe (et française), la BD russe et française. |
MUSEAAV |
|
14h-16h
|
Ateliers du théâtre de Marionnettes de Moscou (fabrication, manipulation) |
MUSEAAV |
|
16h30-17h30
|
Rencontre avec Elana (chants en russe et en français) – dialogue avec le public |
MUSEAAV |
|
15h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La Douce » F.Dostoevski |
Théâtre de la Cité |
|
22 octobre
|
|
|
|
10h -13h-
14h - 18h
|
La presse et les media – internet francorusseRencontres avec journalistes, présentation de la Fondation «le Monde russe » |
MUSEAAV |
|
19h30
|
Concert des lauréats du Concours Véra Lautard (piano). CEDAC de Cimiez en coopération avec l’Association France-Oural. |
CEDAC de Cimiez |
|
23 octobre
|
|
|
|
10h - 13h-
14h – 16h45
|
Traduction et interprétationTraductions officielles, littéraire et poétique ; table ronde : comment devient-on interprète-traducteur. |
MUSEAAV |
|
18h – 20h
|
Rencontre avec Tatyana Danilyants, rencontre poétique. |
MUSEAAV |
|
24 octobre
|
|
|
|
9h - 18h
|
Journée des scolaires |
MUSEAAV |
|
9h30-11h30
|
Ateliers de contes, initiation à la langue russe, marionnettes (premier cycle) |
|
|
14h-16h
|
Alphabets et langues (collège), chants et danses russes |
MUSEAAV |
|
16h-17h30
14h
|
Alphabet, phrases usuelles, échanges sur l’histoire de la Russie, concours, spectacle de marionnettesMarionnettes de Moscou , « La petite oie » |
MUSEAAVThéâtre de la Cité |
| Date |
Contenu |
Lieu |
|
25 octobre
|
|
|
|
14h -18h
|
Rencontreavec les littératures finno-ougriennes et les peuples du nordAvec Eva Toulouze, Sébastien Cagnoli et les acteurs du TNFG |
MUSEAAV |
|
14h30
|
Panorama des littératures finno-ougriennes de Russie en langue française (Komis, Oudmourtes, Vepses…) |
MUSEAAV |
|
15h00
|
Zoom sur les peuples du Nord (vue d’ensemble, cas du poète éleveur de rennes Iouri Vella, lectures de textes…) |
MUSEAAV |
|
16h00
|
Zoom sur les Komis (poésie classique et/ou contemporaine, lecture d’extraits en komi et en français…) |
MUSEAAV |
|
17h00
|
Rencontreavec les acteurs du TNFG (récit de voyage, discussion avec le public)Entre les interventions : présentation-vente de livres. |
MUSEAAV |
|
18h45 – 20h00
|
Concert – Elana (Saratov) |
|
| |
|
|
|
26 octobre
|
|
|
|
9h30 -11h30
11h -17h
|
Accueil des écoles du quartierNissa la Russe. Journée consacrée à Nice et la Côte d’Azur avec les éditeurs locaux, signature des derniers livres sortis sur la présence russe. |
MUSEAAV |
|
18h30
|
Inauguration du monument à Tchékhov |
Hôtel Oasis |
|
21h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La Douce » F.Dostoevski |
Théâtre de la Cité |
|
27 octobre
|
|
|
|
11h – 17h
|
Ateliers du théâtre de Marionnettes de Moscou |
Maison de la Russie |
|
21h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La Douce » F.Dostoevski |
Théâtre de la Cité |
|
28 octobre
|
|
|
|
15h00
|
Spectacle du Theâtre Zertsala (St-Pétersbourg/Lyon).
« La pelisse » N.Gogol |
Théâtre de la Cité |
| 15 – 16 novembre Colloque international – « Guerres et paix. Le choc des idées en Europe au XIXe siècle » |
|
15 novembre
|
|
|
|
9h-12h
14h30-18h30
|
Ouverture officielle et travaux de la première journée «200e anniversaire de Herzen. Exil et Ecriture » |
Théâtre du Grand Château, Valrose |
|
19h00
|
Fin de la journée « Herzen ». Buffet dînatoire pour les participants à la Maison de la Russie à Nice |
Maison de la Russie à Nice |
|
21h00
|
Spectacle « la Barque, la Vague et le Pilote », Simone Rist (Paris) |
Théâtre de la Cité |
|
16 novembre
|
|
|
|
9h-12h
14h30-18h30
|
Ouverture officielle et travaux de la deuxième journée « 200e anniversaire de la bataille de la Moskova. Napoléon 1er et la Russie. » Table ronde. |
Théâtre du Grand Château, Valrose |
|
19h00
|
Clôture du colloque |
|
|
22 novembre
|
|
|
|
19h00
|
Conférence. Les Académies des Saisons Musicales Russes à Nice. |
Eglise Anglicane de Nice |
|
20h00
|
Concert. Point d’Orgue. Gala Russe. Anatoly Safiulin (basse), Regina Shteynman (alto), Philipp Nodel (hautbois), Nestor Nikitin (violon), Elena Keylina (orgue). Bach, Kerll, Abeille, Telemann, Druzhinin |
Eglise Anglicane de Nice |
15 décembre Final du programme «Conte russe sur la Cote d’Azur»
Renseignements et réservations : Maison de la Russie à Nice |
|
Publié le Conférences, Expo Temporaire, Performances |
Publié le Conférences, Expo Temporaire, Performances |




/https%3A%2F%2Fprofilepics.canalblog.com%2Fprofilepics%2F4%2F2%2F426588.jpg)









































































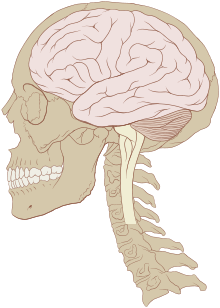
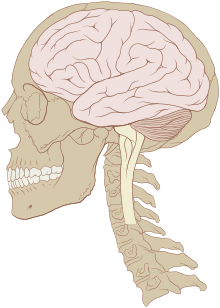
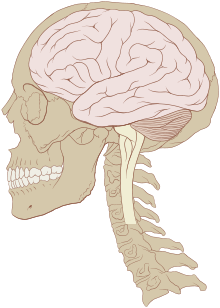

































/http%3A%2F%2Fstorage.canalblog.com%2F72%2F68%2F497307%2F124192998_o.jpg)
/http%3A%2F%2Ffile%3A%2F%2FC%3A%2FUsers%2FGRARD~1%2FAppData%2FLocal%2FIM%2FRuntime%2FSkin%2FE2FEE5~1%2FSTATIC~1.GIF)
/http%3A%2F%2Fstorage.canalblog.com%2F95%2F89%2F497307%2F110069183_o.jpg)
/http%3A%2F%2Fstorage.canalblog.com%2F42%2F39%2F497307%2F107863768_o.jpg)